Vývoj vegetace na opuštěných polích v oblasti Chelčic
The plant communities development on old fields in the area of Chelčice (South Bohemia)
Příspěvek byl uveřejněn ve sborníku
Karel Matějka
Abstract
The plant communities during the 13-year succession of several old fields in the area of border between South Bohemian Basin and Šumava Mts. piedmont is a matter of the paper. Two plots (F and AF) of the fields have been cultivated until 1990 and 1985 respectively. A set of plots of old fields with various management (G1, G2 with a ruderal vegetation, G with a grassland moved till 1986), two stands of Betula pendula on old fields and an oak forest (W) were under comparisons.
Species composition of communities was studied using phytocoenological relevés processed by multivariate analysis methods. Communities were classify into the Braun-Blanquet system, too. A sequence of species Apera spica-venti, Epilobium adenocaulon, Agrostis gigantea, Calamagrostis epigeios was found in first years. Main woody species of the newly abandoned field in the area is Salix caprea. Its development including the biomass production was studied in plot AF.
Changes in vegetation composition are frequently limited by competition (e.g. development of woody species, occurrence of long-term stabile herb communities with Aegopodium podagraria or with some grass species).
A basic characteristic of the vegetation is a biomass production (both above- and below-ground). An important parameter describing changes during succession is the weight relation between live biomass and dead matter.
Dynamics of the trees of the oak (Quercus robur) stand W, of the herb-layer vegetation, and of the organic matter in the plot were compared.
Keywords
classification of vegetation, old field, plant biomass, Quercus robur, Salix caprea, shrub layer development, soil detritus, spatial structure, succession, tree damage
Úvod
Práce se zabývá hodnocením vývoje vegetace na trvalých studijních plochách z oblasti Chelčic v jižních Čechách (okraj Českobudějovické pánve), na nichž byl prováděn od roku 1985 výzkum sukcese (Matějka, 1990). Intensivní sledování ploch bylo přerušeno po roce 1990. Mimo průběžného dalšího zápisu fytocenologických snímků, byl proveden rozbor situace na sledovaných plochách v roce 1997. Samostatně byla hodnocena situace na srovnávací lesní lokalitě (plocha W; viz Matějka, 1994).
Práce byla zpracována v rámci grantu GA ČR reg.č. 526/97/0631 ”Vývoj biodiversity půdního prostředí během sekundární sukcese na opuštěných polích”, jehož řešitelským pracovištěm je Ústav půdní biologie AV ČR v Českých Budějovicích.
Popis ploch
Plocha F
Nejdříve obhospodařované pole (v roce 1986 jarní pšenice, 1987 brambory, 1988 jarní ječmen). V roce 1989 zaseta krmná směska (oves, bob a hrách s podsevem jetele a jílku), ta byla sklizena. V roce následujícím (1990) pokračovala vegetace jetele s jílkem, jejichž kulturu je možno považovat za dvou- až tříletou. V následujícím však již plocha nebyla obhospodařována, začala se vyvíjet jako úhor. Za podstatný moment sukcese je nutno považovat způsob opuštění, kdy nenastala etapa, kdy půda není kryta vegetací (jak tomu bylo na následující ploše AF na přelomu let 1985 a 1986). Již od začátku vývoje převažovaly víceleté plevele, jejich zastoupení se až do roku 1997 měnilo plynule, postupně se ve struktuře společenstva zvětšovala účast lučních druhů. V poslední době přistupují rovněž druhy charakteristické pro paseky. Přitom dosud nelze mluvit o významnější eliminaci některých druhů tvořících společentvo na počátku sukcese. Oba jevy mají efekt v postupně se zvyšující druhové diversitě.
Plocha AF
Úhor vznikl na konci léta 1985, kdy byla sklizena poslední plodina - krmná kukuřice. Po sklizni byla provedena podmítka. Vzhledem k aplikaci triazinových herbicidů při setí kukuřice byl zaznamenán silnější rozvoj vegetace až v roce následujícím. Vývoj na této ploše lze rozdělit do čtyř etap:
1. etapa (rok 1986) s druhy jednoletých plevelů s převahou Apera spica-venti
2. etapa (1987-1989) charakterizovaná druhy Epilobium adenocaulon a Cirsium arvense
3. etapa (1990-1994) s výraznou dominantou Calamagrostis epigeios
4. etapa (od přelomu let 1994 a 1995 dosud), kdy se výrazně vyvinulo keřové patro se Salix caprea
Přechod mezi druhou a třetí etapou byl plynulý. Rovněž přechod mezi etapou třetí a čtvrtou nebyl náhlý, ale jeho základ lze spatřovat již v postupném zvyšování účasti vrby v předchozím období v rámci bylinného patra. Stav v roce 1997 je charakteristický velmi hustým keřovým patrem s výškou až 4 m působícím nedostatek světla v podrostu, který je relativně druhově bohatý, pouze málo druhů je významnějších, bylinná etáž má nízkou celkovou biomasu a většina druhů není fertilních. Vzhledem k nízké ploše společetstva vyvinutého v pásu širokém cca 15 m je nutno vždy brát v úvahu fakt ovlivnění plochy ekotonálním efektem, který je výraznější se vzrůstající výškou keřů.
Plocha G1
Tato plocha sousedí s plochami AF a W0. Jedná se o starý třešňový sad s dvěma řadami stromů, jehož střed byl dříve orán a obhospodařován jako pole. Obhospodařování bylo ukončeno přibližně v polovině sedmdesátých let. Dominantami v roce 1985 byly Agropyron repens, Aegopodium podagraria, Urtica dioica a Cirsium arvense. Srovnání skladby společenstva bylo provedeno jednorázově po dvanácti letech. Vývoj na lokalitě byl velmi pomalý, dominanty se prakticky nezměnily, rovněž skladba doprovodných druhů je obdobná. To vyvolává otázku, proč je dané společestvo s Aegopodium podagraria tak stabilní. Pravděpodobně se jedná o tak konkurenčně silný druh, který změnu nedovolí. To je fakt, který je známý i z jiných stanovišť a jenž by si jistě zasloužil širší pozornost.
Plocha G
Lokalita se nachází v blízkosti ploch AF, G1 a W0. Jde o v minulosti běžně obhospodařované pole opuštěné okolo roku 1973. Do roku 1984 byla celá plocha pravidelně kosena, v roce 1985 byla pokosena pouze severní část lokality. Oproti předcházejícím plochám umístěným na mírném svahu, tato je lokalisována v široké sedlovité terénní depresi, kde hladina spodní vody zasahuje po větší část roku k silně prokořenělým povrchovým půdním horizontům (tento fakt se začal projevovat až následně po vyloučení obhospodařování v souvislosti s postupně se snižující funkčností melioračních objektů). Společenstvo na lokalitě je poměrně stabiln@?. Eliminací kosení došlo ihned v prvních letech k velmi významnému ústupu dvouděložných lučních druhů, čímž se snížila ”optická diversita”. Porovnáním stavu vegetace v roce 1997 s předchozím rozdělením tří základních společenstev zjištěných v roce 1988, je možno konstatovat stabilitu plošného rozdělení vegetačních typů. Přestože nebylo provedeno přesné sledování, zdá se, že největší změny ve výše uvedeném smyslu proběhly ve společenstvu Hypericum maculatum-Arrhenatherum elatius, nejnižší pak pravděpodobně v nejvlhčí části lokality (společenstvo Cirsium palustre-Juncus effusus).
Plocha G2
Jedná se o soustavu stupňovitých teras s úhorovými porosty. Hospodaření na lokalitě bylo ukončeno asi v druhé polovině sedmdesátých let. Na lokalitě s účastí řady teplomilných druhů je možno sledovat serii ekotonálních efektů.Opuštění teras neproběhlo ve stejnou dobu. Tím byla způsobena rozdílnost jednotlivých částí lokality. Nový fytocenologický zápis byl proveden v horní části první terasy, která se jevila mít sukcesně nejmladší rostlinné společenstvo. Zaznamenané desetileté změny jsou poměrně malé, za podstatné lze považovat úplný ústup Agropyron repens a Galium aparine, naopak rozvoj Arrhenatherum elatius, Selinum carvifolia a druhů teplomilných lemů (např. Verbascum nigrum a Centaurea scabiosa).
Plocha B1
Starší úhor s porostem Betula pendula. Plocha navazuje na les a je na svém jihovýchodním okraji v kontaktu s obhospodařovaným polem. Stáří se v současnosti odhaduje na 45 až 50 let.
Plocha B2
Srovnávací lokalita obdobná předcházející, která byla hodnocena z hlediska fytocenologického na začátku výzkumu.
Plocha W
Kulturní listnatý les s více méně přirozeným druhovým složením. Věková skladba porostu však neodpovídá stavu přirozených porostů. Vývoj této plochy byl sledován samostatně a přehled výsledků byl podán již dříve (Matějka, 1994).
Plocha W0
Kulturní smrkový porost, který se nachází v kontaktu ploch F, AF, G1 a G. Díky tomu se může stát i zvlášť významným zdrojem diaspor lesních druhů, které se potenciálně mohou uplatňovat v průběhu sukcese na těchto lokalitách..
Analýza vývoje vegetace
V různých časových intervalech byly zapisovány klasické fytocenologické snímky. Pro účely této práce bylo hodnocení vývoje vegetace na jednotlivých plochách a vztahy mezi rostlinnými společenstvy těchto ploch hodnoceny na základě klasifikace (hierarchická aglomerativní klasifikace: Wardova metoda a average lincage) a ordinace (metoda DCA) bylinné etáže.
Obě použité klasifikační metody demonstrují velký vliv použité metody na konečné výsledky a nutnost jejich uvážení při konečné interpretaci. Vegetace vykazovala velké rozdíly mezi jednotlivými plochami i na téže ploše v průběhu sukcese. Zajímavá je poměrně velká podobnost mezi některými sukcesně mladými (i 5 až 10 let strarými) porosty a srovnávanými lesními společenstvy. Naopak značně odlišně se jeví iniciální stadia sukcese (společenstva jednoletých plevelů).
Přehled definovaných vegetačních jednotek (v závorce jsou uvedeny plocha, případně rok výskytu na lokalitě; všechna zaznamenaná společenstva nejsou klasifikována v rámci následujícího přehledu):
třída Secalietea
řád Aperetalia
svaz Aphanion
společenstvo Apera spica-venti (AF: 1986)
třída Galio-Urticetea
bas. spol. Urtica dioica-Aegopodium podagraria-[Galio-Urticetea] (G1)
bas. spol. Urtica dioica-[Galio-Urticetea] (část plochy G, 1985)
odvoz. spol. Agropyron repens-[Galio-Urticetea] (G1)
třída Agropyretea repentis
řád Agropyretalia repentis
svaz Convolvulo-Agropyrion
odvoz. spol. Agropyron repens-Hypericum perforatum-[Galio-Urticetea/Agropyretea] (G2)
společenstvo Epilobium adenocaulon-Agrostis gigantea (AF: 1987-1989)
společenstvo Calamagrostis epigeios (AF: 1990-1994)
třída Epilobietea angustifolii
řád Sambucetalia
svaz Sambuco-Salicion capreae
společenstvo Salix caprea (AF: od 1995)
třída Molinio-Arrhenatheretea
řád Arrhenatheretalia
svaz Arrhenatherion
odvoz. spol. Agropyron repens-Holcus mollis-[Arrhenatherion] (G)
třída Quercetea robori-petraeae
řád Quercetalia robori-petraeae
svaz Genisto germanicae-Quercion
společenstvo s Betula pendula (B1, B2)
třída Querco-Fagetea
řád Fagetalia sylvaticae
svaz Fagion
asociace Abieto-Quercetum (W)
Nadzemní rostlinná biomasa
Rostlinná biomasa byla odstříhávána při půdním povrchu z ploch velikosti 0,25 m2 a následně veškerý detrit byl z tohoto půdního povrchu sebrán. Rozdělení odebrané hmoty na živou a mrtvou (dále označovanou jako detrit, ve skutečnosti detrit a standing dead matter), případně na biomasu jednotlivých druhů, byly provedeny v laboratoři. Jednotlivé vzorky byly sušeny při 80 C do konstantní hmotnosti. Poslední odběr byl proveden 22. 7. 1997.
Rozbor keřové biomasy na ploše AF je uveden ve zvláštní kapitole, stromová biomasa na ploše W byla hodnocena dříve (Matějka, 1994).
Získané výsledky lze interpretovat následujícím způsobem:
Podzemní rostlinná biomasa a půdní detrit
Podzemní biomasa a půdní detrit byly sledovány na základě odebraných půdních monolitů odebíraných kruhovou sondou o průměru 65 mm. Standardní počet opakování na každé ploše byl 5. Monolity byly odebírány z hloubky 0-5 cm a 5-10 cm (jiný způsob byl praktikován na ploše W, celková hloubka pro odběr byla 10 cm).Vzorky byly zmraženy pro narušení kompaktnosti půdy. Po rozmražení byly rozplavovány ve vodě a promývány proudem vody na sítě s průměrem ok 0,25 mm, přičemž byla vybírána veškerá podzemní rostlinná biomasa. Zbytek vzorku byl sušen při 80 C a z usušeného vzorku byla vodou vyplavována zbylá organická hmota (takzvaný půdní detrit).
Poměr živé a mrtvé podzemní biomasy byl stanoven za pomoci barvení Kongo-červení (viz např. Tesařová, Fiala, Studený, 1982) na základě speciálně odebraných malých půdních monolitů o ploše cca 4 cm2. Kořenová hmota byla vybírána v průběhu rozplavování monolitů proudem vody (vzorky nebyly předem mraženy).
Získané výsledky je možno hodnotit následovně:
Keřová etáž na ploše AF
Vzhledem k zaznamenanému mohutnému rozvoji keřové vegetace na ploše AF, byla v roce 1997 pozornost věnována keřové dominantě, jíž se stala Salix caprea. Cílem bylo
Pro tento cíl byly zaznamenány všechny keře S. caprea na třech zkusných ploškách velikosti 2m × 2m s tím, že u každého jedince byl zaznamenán jeho bazální průměr (průměr ve výšce cca 10 cm). Odebráno bylo pět vzorníků keřů různých velikostních kategorií, u nichž byly změřeny parametry bazální průměr ve výšce 10 cm, biomasa letorostů (zvlášť listy a roční přírůsty větví) a biomasa dřeva. U čtyř největších vzorníků byl analyzován průřez bazí a byla provedena jednoduchá letokruhová analýza při níž byla změřena plocha jednotlivých ročních přírůstů a ta byla dále zpracovávána
Z uvedených hodnot lze provést následující závěry:
Na základě zjištěných hmot jednotlivých částí biomasy vzorníků vrb (tab. 10) byly zjištěny následující regresní vztahy:
log(L) = 1.0223 + 1.2767 log(BA) |
r = 0.997 |
log(S) = 0.5605 + 1.2538 log(BA) |
r = 0.979 |
log(W) = 1.7134 + 1.3142 log(BA) |
r = 0.998 |
Všechny vztahy jsou vysoce průkazné (n=4, vzorníky 3 a 4 byly pro svou podobnost nahraženy jejich průměrem). Na základě těchto alometrických vztahů a výskytu jedinců vrb o známých kruhových základnách na třech plochách (ve čtvercích 4, 8 a 12) velikosti 4 m2 byl vypočten odhad biomasy na jednotku plochy (tab. 11).
Dále byl proveden pokus o odhad přírůstu celkové nadzemní biomasy bez letorostů (tzv. biomasa dřeva) na základě výsledků letokruhové analýzy a zjištěného alometrického modelu. Vypočtené zásoby na konci jednotlivých let byly:
1992 |
483 g.m-2 |
1993 |
575 g.m-2 |
1994 |
776 g.m-2 |
1995 |
1022 g.m-2 |
1996 |
1368 g.m-2 |
Vzhledem k zjištěnému poměru mezi biomasou letorostů s listy k biomase dřeva ve výši přibližně 0,25:1 , lze provést i odhad roční produkce opadu ve výši asi jedné čtvrtiny uvedených zásob biomasy dřeva.
Stromové patro na lokalitě W
Základní údaje o porostu je možno získat z lesního hospodářského plánu - ten byl vypracován s platností pro období 1988-1997. Porost je označen jako 74B2. Uvedený lesní typ 3S1 (svěží dubová bučina šťavelová), hospodářský soubor 255, zakmenění 9, věk (k roku 1997) 68 let. Charakteristiky jednotlivých dřevin dle LHP jsou následující:
druh |
zastoupení [%] |
DBH [cm] |
střední výška [m] |
zásoba [m3/ha] |
dub |
60 |
22 |
21 |
131 |
smrk |
25 |
20 |
20 |
80 |
lípa |
10 |
20 |
19 |
20 |
buk |
5 |
20 |
19 |
10 |
Všechny stromy byly zaměřeny na ploše velikosti 30m× 15m. Opakovaně byly hodnoceny dendrometrické parametry a defoliace. V úvahu jsou dále rovněž vzaty výsledky provedené letokruhové analýzy.
Výsledky lze interpretovat následovně:
Závěr
Literatura
delMoral R., Wood D.M. (1988): Dynamics of herbaceous vegetation recovery on Mount St. Helens, Washington, USA, after a volcanic eruption. - Vegetatio, 74: 11-27.
delMoral R., Wood D.M. (1993): Early Primary Succession on the Volcano Mount St-Helens. - J. Veg. Sci., 4: 223-234.
Halaj J., Grék J., Pánek F., Petráš R., Řehák J. (1987): Rastové tabuľky hlavných drevín ČSSR. - Bratislava: 361pp.
Kellman M., Kading M. (1992): Facilitation of Tree Seedling Establishment in a Sand Dune Succession. - J. Veg. Sci., 3: 679-688.
Matějka K. (1990): Struktura a funkce společenstev primárních producentů ve vztahu k půdní složce ekosystémů. - Ms. [Kandidátská disertační práce, ÚPB České Budějovice]
Matějka K. (1994): Several results of the study in oak forest in periphery of the South Bohemian Basin. In K. Matějka, ed., Investigation of the forest ecosystems and of forest damage. Lowland and submountain forests and monitoring of the forests status. - Praha, p. 74-92.
McCook L.J. (1994): Understanding Ecological Community Succession - Causal Models and Theories, a Review. - Vegetatio, 110: 115-147.
Tesařová M., Fiala K., Studený V. (1982): Live and dead roots - their mass ratio in several grassland stands. - Folia Geobot. Phytotax., 17: 127-130.
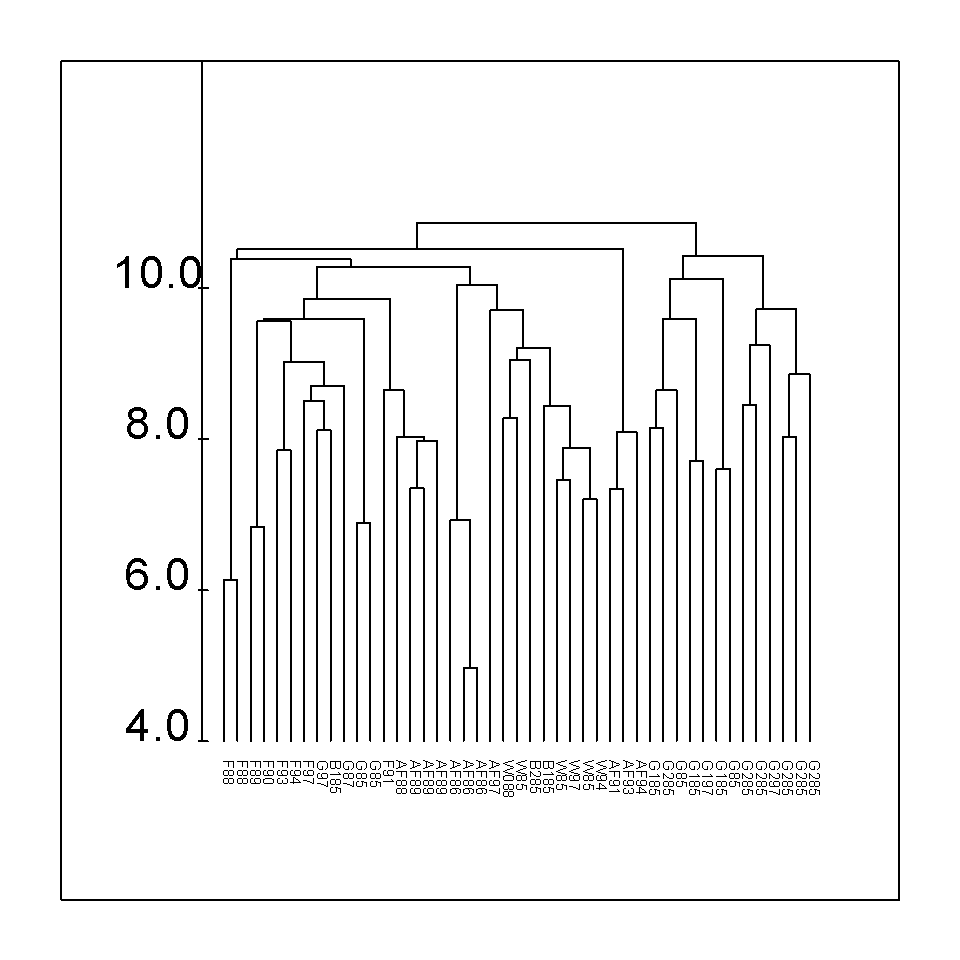
Obr. 1: Klasifikace bylinné etáže dle fytocenologických snímků na studovaných plochách v oblasti Chelčic. Použita Wardova hierarchická klasifikační metoda, kvadrát euklidovské distance jako míra nepodobnosti; osa Y je zobrazena v logaritmickém měřítku.
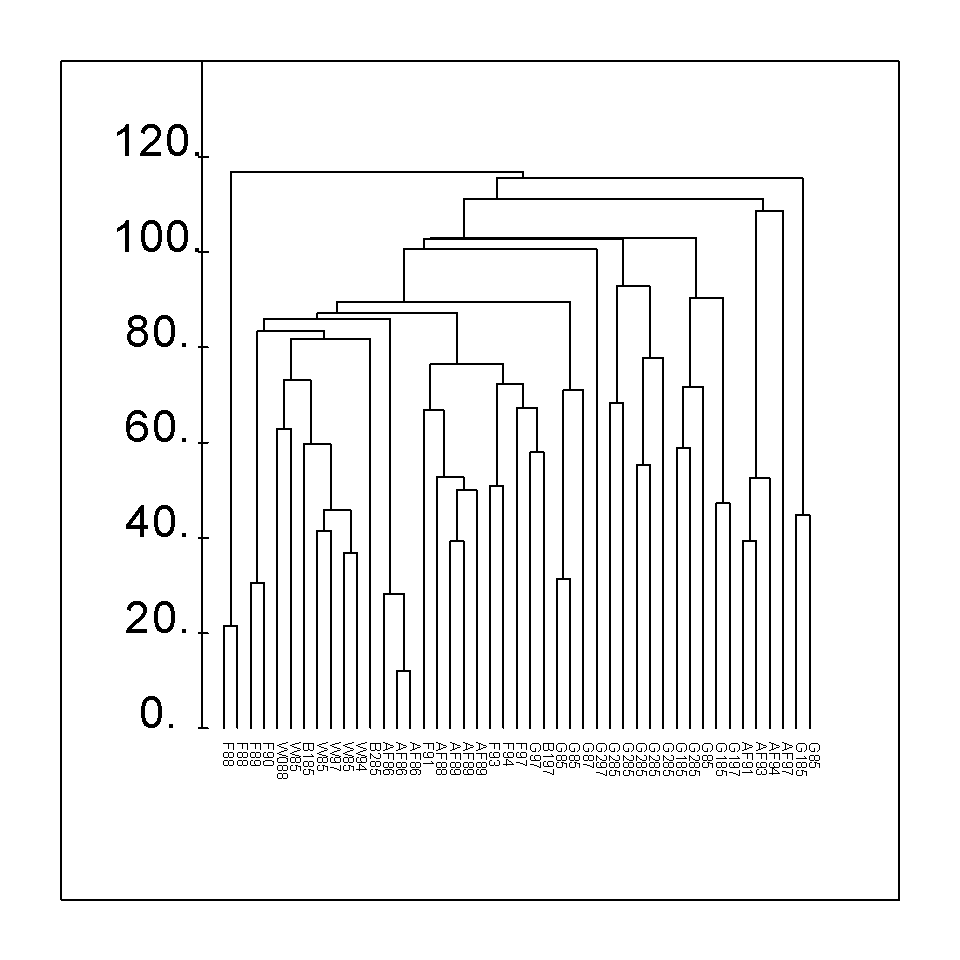
Obr. 2: Klasifikace bylinné etáže dle fytocenologických snímků na studovaných plochách v oblasti Chelčic. Použita metoda ”average distance” hierarchické klasifikace, euklidovská distance jako míra nepodobnosti.
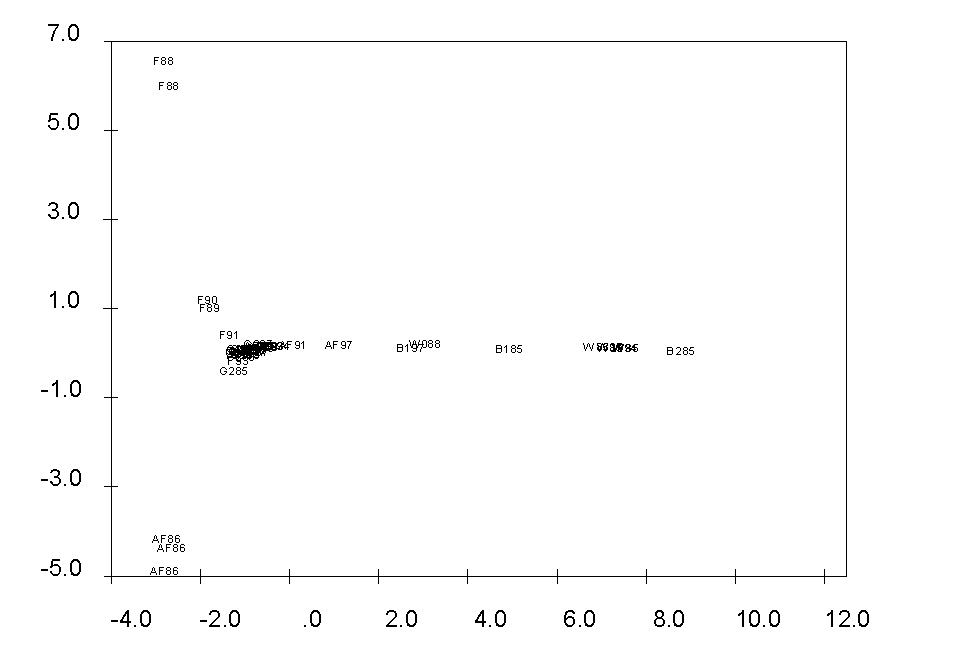
Obr. 3: Ordinace všech fytocenologických snímků metodou DCA (osa I a II). Snímky označeny plochou a dvoučíslím roku sledování.
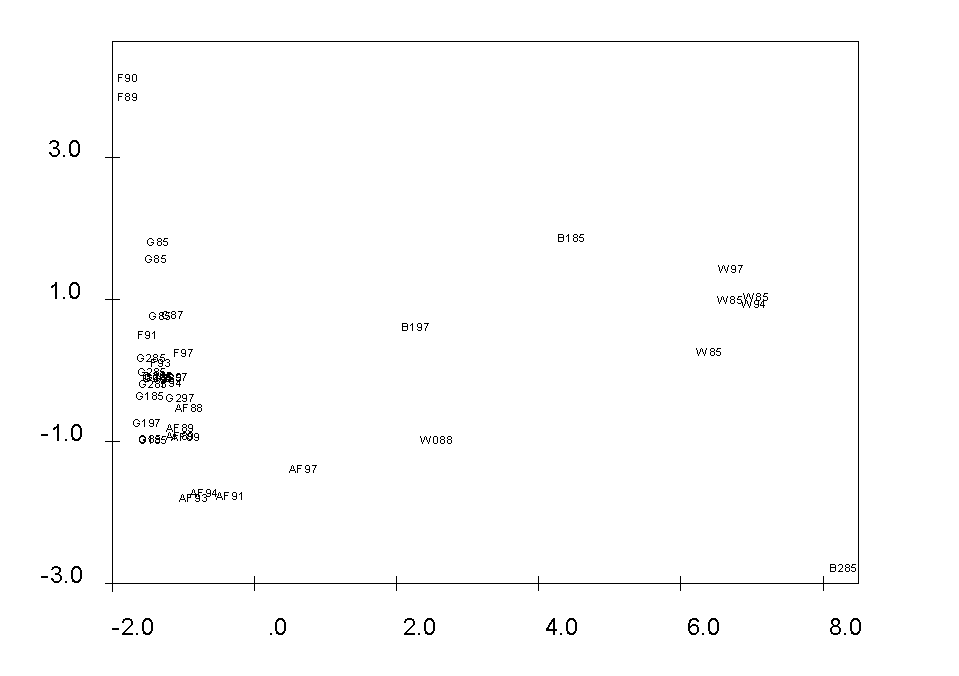
Obr. 4: Ordinace fytocenologických snímků s vyloučením iniciálních společenstev sukcese (metoda DCA, osy I a II)
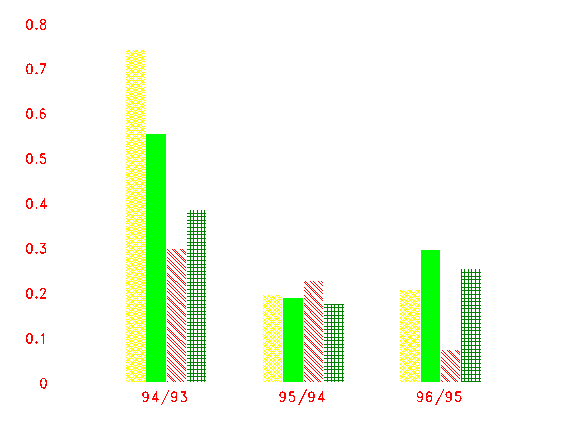
Obr. 5: Index přírůstu Salix caprea pro tři období počítané pro čtyři vzorníky (postupně 1 až 4).
index = log (P1/P0) P1, P0 - plocha letokruhu aktuálního resp. předcházejícího roku
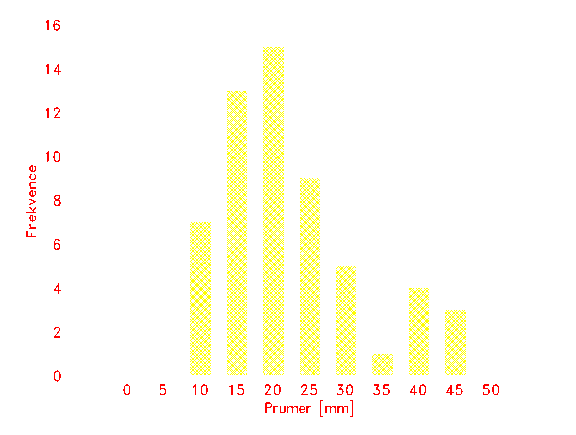
Obr. 6: Frekvence jedinců Salix caprea ve třech kvadrátech velikosti 4 m2 na lokalitě AF dle jejich bazálního průměru.
Tab. 1: Nadzemní biomasa a její složení na ploše F v roce 1997. Údaje u jednotlivých vzorků jsou v gramech na vzorek (1/4 m2).
3 |
4 |
8 |
12 |
celkem na 1 m2 |
||
Achillea millefolium |
23.96 |
7.58 |
3.75 |
5.73 |
41.02 |
|
Aegopodium podagraria |
3.11 |
3.11 |
||||
Cirsium arvense |
8.30 |
8.30 |
||||
Crepis biennis |
+ |
|||||
Mentha arvensis |
+ |
+ |
+ |
|||
Plantago uliginosa |
+ |
|||||
Ranunculus repens |
+ |
+ |
+ |
+ |
||
Rumex obtusifolius |
+ |
|||||
Sonchus arvensis |
+ |
8.33 |
8.33 |
|||
Stachys palustris |
+ |
|||||
Stellaria graminea |
+ |
+ |
+ |
|||
Taraxacum officinale agg. |
+ |
+ |
+ |
+ |
||
Urtica dioica |
3.34 |
6.52 |
9.85 |
|||
Vicia hirsuta+ V.angustifolia (8) |
11.89 |
2.29 |
4.83 |
+ |
19.01 |
|
ostatní dvouděložné |
18.50 |
5.72 |
3.72 |
4.24 |
||
trávy celkem |
47.59 |
60.20 |
101.61 |
56.58 |
265.99 |
|
detrit |
39.71 |
27.81 |
68.14 |
52.25 |
187.90 |
|
živá biomasa celkem |
101.95 |
90.58 |
113.92 |
81.37 |
387.81 |
Tab. 2: Nadzemní biomasa a její složení na ploše AF v roce 1997. Údaje u jednotlivých vzorků jsou v gramech na vzorek (1/4 m2).
3 |
4 |
8 |
12 |
15 |
celkem na 1 m2 |
|
Achillea millefolium |
0.08 |
0.11 |
0.15 |
0.27 |
||
Equisetum sylvaticum |
+ |
|||||
Fragaria sp.div. |
0.69 |
0.20 |
0.72 |
|||
Galium aparine |
+ |
|||||
Hypericum perforatum |
0.83 |
0.67 |
||||
Selinum carvifolia |
0.42 |
0.27 |
0.55 |
|||
Trifolium pratense |
1.70 |
1.36 |
||||
Urtica dioica |
1.04 |
+ |
0.81 |
1.48 |
||
Vicia hirsuta |
0.19 |
+ |
0.16 |
0.27 |
0.15 |
0.62 |
ostatní dvouděložné |
0.25 |
0.67 |
0.13 |
0.04 |
0.10 |
|
trávy |
16.85 |
6.40 |
6.60 |
10.27 |
19.37 |
47.58 |
detrit |
62.62 |
65.90 |
98.03 |
81.00 |
71.84 |
303.51 |
živá biomasa celkem |
18.33 |
7.49 |
7.93 |
14.02 |
19.97 |
54.20 |
Tab. 3: Nadzemní biomasa na ploše G v roce 1997. Údaje u jednotlivých vzorků jsou v gramech na vzorek (1/4 m2).
3 |
4 |
8 |
12 |
15 |
celkem na 1 m2 |
|
celková nadzemní biomasa |
168.35 |
239.51 |
209.09 |
237.50 |
213.16 |
854.09 |
Tab. 4: Nadzemní biomasa a její složení na ploše W v roce 1997. Údaje u jednotlivých vzorků jsou v gramech na vzorek (1/4 m2).
3 |
4 |
8 |
12 |
15 |
celkem na 1 m2 |
|
Calamagrostis arundinacea |
20.88 |
16.70 |
||||
Fragaria sp.div. |
6.95 |
1.30 |
2.16 |
8.33 |
||
Fraxinus excelsior juv. |
2.71 |
2.17 |
||||
Galium aparine |
0.59 |
0.48 |
||||
Galium rotundifolium |
0.49 |
0.14 |
0.54 |
0.93 |
||
Geum urbanum |
1.89 |
1.51 |
||||
Impatiens parviflora |
0.24 |
0.13 |
0.30 |
|||
Melampyrum pratense |
0.61 |
2.97 |
0.20 |
1.62 |
4.32 |
|
Moehringia trinerviva |
+ |
0.32 |
0.26 |
|||
Mycelis muralis |
0.37 |
0.30 |
||||
Oxalis acetosella |
0.38 |
0.68 |
3.23 |
1.91 |
2.82 |
7.21 |
Poa nemoralis |
34.00 |
27.20 |
||||
Sanicula europaea |
5.21 |
4.14 |
6.10 |
0.76 |
12.97 |
|
Senecio fuchsii |
3.51 |
2.81 |
||||
Vaccinium myrtillus |
5.07 |
10.75 |
7.37 |
18.55 |
||
Veronica chamaedrys |
0.93 |
9.55 |
8.51 |
15.19 |
||
Viola reichenbachiana |
0.87 |
0.59 |
1.41 |
2.30 |
||
ostatní dvouděložné |
0.06 |
0.05 |
||||
trávy celkem |
2.66 |
* |
3.71 |
* |
0.40 |
|
detrit |
81.90 |
66.41 |
42.51 |
91.65 |
74.94 |
285.93 |
živá biomasa celkem |
19.77 |
39.621 |
35.237 |
35.379 |
28.777 |
127.03 |
Poznámka: Vzorek 4 náleží k typu mikrofytocenózy Poa nemoralis, vzorek 12 k t.mf. Calamagrostis arundinacea, ostatní vzorky lze charakterizovat pravděpodobně jako t.mf. Luzula-Veronica chamaedrys.
Tab. 5: Souhrnné údaje o nadzemní biomase na plochách v průběhu sukcese (údaje v g.m-2).
plocha |
datum |
počet vzorků |
velikost (m2) |
nadzemní biomasa [g.m-2] |
|||
|
|
|
|
D |
L |
T |
L/D |
F |
7. 6. 1989 |
2 |
1/4 |
|
338.5 |
338.5 |
|
|
22. 7. 1997 |
4 |
1/4 |
187.9 |
387.8 |
575.7 |
2.06 |
AF |
5. 8. 1986 |
4 |
1/4 |
18.8 |
267.1 |
285.8 |
14.20 |
|
21. 4. 1987 |
|
1/4 |
156.0 |
|
|
|
|
7. 7. |
5 |
1/4 |
mimo sled. plochy |
344.0 |
|
|
|
8. 7. |
|
1/16 |
181.1 |
251.0 |
432.1 |
1.39 |
|
6. 8. |
5 |
1/4 |
204.8 |
316.4 |
521.2 |
1.54 |
|
5. 10. |
4 |
1/16 |
233.5 |
286.8 |
520.2 |
1.23 |
|
20. 4. 1988 |
5 |
1/16 |
306.7 |
52.5 |
359.2 |
.17 |
|
25. 5. |
5 |
1/16 |
212.7 |
181.2 |
393.9 |
.85 |
|
20. 6. |
20 |
1/16 |
221.4 |
333.8 |
555.2 |
1.51 |
|
5. 9. |
5 |
1/4 |
332.8 |
261.2 |
594.0 |
.52 |
|
14. 3. 1989 |
5 |
1/8 |
754.6 |
|
|
|
|
10. 5. |
5 |
1/8 |
584.3 |
97.4 |
681.7 |
.17 |
|
17. 7. |
5 |
1/8 |
477.5 |
342.6 |
820.1 |
.72 |
|
9.10. |
5 |
1/8 |
641.7 |
191.4 |
833.1 |
.30 |
|
22. 7. 1997 |
5 |
1/4 |
|
54.2 |
|
0.18 |
|
-Salix caprea (listy) |
|
|
323.5 |
|
|
|
|
-celkem |
|
303.5 |
377.7 |
681.2 |
1.24 |
|
G |
8. 8. 1985 |
5 1/4 |
|
|
|
531.5 |
11.3 |
|
19. 8. 1986 |
5 1/4 |
|
|
|
434.6 |
|
|
8. 7. 1987 |
|
1/16 |
271.4 |
626.8 |
898.2 |
2.31 |
|
19. 8. |
|
1/4 |
|
|
627.5 |
|
|
12. 10. |
2 |
1/16 |
357.2 |
163.1 |
520.3 |
.46 |
|
20. 4. 1988 |
5 |
1/16 |
|
|
478.8 |
|
|
25. 5. |
5 |
1/16 |
502.0 |
159.3 |
661.3 |
.32 |
|
20. 6. |
16 |
1/16 |
317.7 |
464.7 |
782.4 |
1.46 |
|
5. 9. |
4 |
1/4 |
|
|
709.4 |
.95 |
|
14. 3. 1989 |
5 |
1/8 |
|
|
593.0 |
|
|
10. 5. |
5 |
1/8 |
442.4 |
155.9 |
598.3 |
.35 |
|
17. 7. |
5 |
1/8 |
419.5 |
588.5 |
1008.0 |
1.40 |
|
9. 10. |
5 |
1/8 |
|
|
633.6 |
|
|
22. 7. 1997 |
5 |
1/4 |
|
|
854.1 |
|
W |
5. 8. 1986 |
6 |
1/4 |
|
54.4 |
|
|
|
8. 7. 1987 |
|
1/16 |
442.8 |
|
|
|
|
6. 8. |
5 |
1/4 |
|
86.4 |
|
|
|
12. 10. |
5 |
1/16 |
422.5 |
60.8 |
|
|
|
20. 4. 1988 |
5 |
1/16 |
387.4 |
|
|
|
|
20. 6. |
5 |
1/4 |
579.3 |
105.1 |
|
|
|
14. 3. 1989 |
5 |
1/4 |
456.9 |
|
|
|
|
3. 6. |
4 |
1/4 |
313.5 |
|
|
|
|
15. 9. |
5 |
1/4 |
254.9 |
155.8 |
|
|
|
22. 7. 1997 |
5 |
1/4 |
285.9 |
127.0 |
|
|
Tab. 6: Poměr živé a mrtvé podzemní biomasy (L/D ratio) vyjádřený jako log(L/D) na lokalitách F, AF a G.
kvadrát: |
3 |
4 |
8 |
12 |
15 |
průměr |
sx |
L/D |
plocha F: |
|
|
|
|
|
|
|
|
8. 7. 1997 |
.07 |
.36 |
.47 |
.55 |
.32 |
.36 |
.16 |
2.26 |
plocha AF: |
|
|
|
|
|
|
|
|
4. 4. 1989 |
.12 |
.77 |
.42 |
-.47 |
.76 |
.32 |
.46 |
2.08 |
7. 6. 1989 |
.52 |
1.18 |
.59 |
-.10 |
.41 |
.52 |
.41 |
3.30 |
5. 9. 1989 |
.68 |
.16 |
.71 |
-.29 |
-.45 |
.16 |
.48 |
1.45 |
16. 11. 1989 |
1.25 |
.79 |
1.26 |
1.09 |
.35 |
.95 |
.34 |
8.87 |
8. 7. 1997 |
.42 |
-1.13 |
.09 |
-.67 |
-.16 |
-.29 |
.55 |
.51 |
plocha G: |
|
|
|
|
|
|
|
|
4. 4. 1989 |
.54 |
.09 |
.54 |
1.10 |
.57 |
.36 |
3.70 |
|
7. 6. 1989 |
-.24 |
-.03 |
.32 |
.14 |
.73 |
.18 |
.33 |
1.53 |
5. 9. 1989 |
-.04 |
.41 |
1.07 |
-.84 |
.95 |
.31 |
.70 |
2.05 |
16. 11. 1989 |
.19 |
.36 |
.20 |
-.27 |
.42 |
.18 |
.24 |
1.51 |
8. 7.1997 |
.18 |
.77 |
-.79 |
-.35 |
-1.30 |
-.30 |
.72 |
.50 |
Tab. 7a: Zásoba celkové podzemní biomasy a půdního detritu na plochách F, G a W. Primární údaje jsou v gramech na vzorek (odebíraný kruhovou sondou o průměru 65 mm).
Podzemní biomasa |
Půdní detrit |
|||||
F |
G |
W |
F |
G |
W |
|
vrstva A - 0-5cm |
||||||
3 |
2.456 |
1.353 |
0.913 |
2.133 |
6.634 |
1.650 |
4 |
3.372 |
0.222 |
1.076 |
2.145 |
3.752 |
3.699 |
8 |
1.699 |
2.189 |
1.679 |
1.762 |
3.340 |
15.800 |
12 |
1.998 |
3.707 |
1.296 |
1.527 |
4.045 |
14.925 |
15 |
2.632 |
3.255 |
1.278 |
8.638 |
5.175 |
16.945 |
celkem [g.m-2] |
732.7 |
646.5 |
376.2 |
976.7 |
1383.0 |
3195.6 |
vrstva B - 5-10cm |
||||||
3 |
0.287 |
0.112 |
0.198 |
0.515 |
0.542 |
0.855 |
4 |
1.604 |
0.068 |
0.195 |
0.490 |
0.622 |
0.695 |
8 |
1.557 |
1.020 |
0.343 |
1.114 |
1.330 |
5.847 |
12 |
0.589 |
1.113 |
0.199 |
0.688 |
1.052 |
7.371 |
15 |
0.705 |
0.906 |
0.469 |
5.590 |
2.409 |
16.034 |
celkem [g.m-2] |
285.8 |
194.0 |
84.6 |
506.1 |
358.9 |
1856.5 |
A+B [g.m-2] |
1018.5 |
840.5 |
460.8 |
1482.8 |
1741.9 |
5052.1 |
poměr A/B |
2.56 |
3.33 |
4.45 |
1.93 |
3.85 |
1.72 |
Tab. 7b: Zásoba celkové podzemní biomasy a půdního detritu na ploše AF. Primární údaje jsou v gramech na vzorek (odebíraný kruhovou sondou o průměru 65 mm).
hloubka [cm] |
podzem. biomasa |
detrit |
|
3 |
10 |
2.158 |
2.847 |
4 |
9 |
2.176 |
2.193 |
8 |
11 |
2.278 |
3.106 |
12 |
11 |
2.745 |
4.587 |
15 |
9 |
3.150 |
1.410 |
celkem [g.m-2] |
753.8 |
852.4 |
|
Tab. 8: Zjištěné zásoby půdního detritu a podzemní biomasy v průběhu sukcese (údaje v g.m-2).
A. půdní detrit
rok |
1987 |
1988 |
1989 |
1997 |
F |
690 |
260-1030 |
240-630 |
1483 |
AF |
650 |
410-1410 |
250-630 |
852 |
G |
1030 |
880-2700 |
640-970 |
1742 |
W |
2290 |
2360-6350 |
1380-2970 |
5052 |
B. podzemní biomasa
rok |
1987 |
1988 |
1989 |
1997 |
F |
0-120 |
10-160 |
1019 |
|
AF |
320 |
180-600 |
200-570 |
754 |
G |
1840 |
860-2280 |
770-1190 |
840 |
W |
550 |
470-790 |
520-660 |
461 |
Tab. 9: Průměrné zjištěné šířky letokruhů a celkové průměry jednotlivých analyzovaných vzorníků Salix caprea (všechny údaje v mm).
vzorník číslo |
1 |
2 |
3 |
4 |
kůra |
1.58 |
1.31 |
1.41 |
1.50 |
přírůst 1997 (*) |
3.23 |
2.94 |
0.99 |
1.56 |
1996 |
5.77 |
3.27 |
1.68 |
2.26 |
1995 |
5.09 |
2.36 |
1.85 |
1.72 |
1994 |
5.48 |
2.37 |
1.57 |
1.66 |
1993 |
1.89 |
1.12 |
1.19 |
1.06 |
průměr celkem |
52.33 |
30.63 |
21.64 |
23.34 |
průměr dle plochy |
52.55 |
30.79 |
21.76 |
23.46 |
(*) je nutno poznamenat, že růst v roce 1997 ještě nebyl ukončen v době odběru.
Tab. 10: Biomasa vzorníků Salix caprea.
vzorník |
BA: kruhová základna (cm2) |
L: listová biomasa (g) |
S: biomasa větví letorostů (g) |
W: hmota dřeva (*) (g) |
1 |
22.06 |
502.02 |
160.00 |
3205.97 |
2 |
7.35 |
158.90 |
62.80 |
681.10 |
3,4 (průměr) |
3.78 |
55.37 |
13.83 |
265.22 |
5 |
1.61 |
18.48 |
7.13 |
106.15 |
(*) Zahrnuje veškerou nadzemní biomasu mimo letorostů s listy; keře odřezány ve výšce cca 5 cm nad povrchem země.
Tab. 11: Přepočet biomasy Salix caprea na jednotkovou plochu ve třech zkusných kvadrátech.
plocha |
BA: kruhová základna (cm2/m2) |
L: listová biomasa (g/m2) |
S: biomasa větví letorostů (g/m2) |
W: hmota dřeva (g/m2) |
4 |
9.15 |
178.5 |
58.5 |
954 |
8 |
23.48 |
366.0 |
122.1 |
1902 |
12 |
22.89 |
426.1 |
140.1 |
2267 |
průměr |
18.51 |
323.5 |
106.9 |
1708 |
Tab. 12: Charakteristiky jednotlivých stromů na ploše v průběhu sledování.
Druh: Q - Quercus robur, A - Acer pseudoplatanus; Výška v metrech; DBH - průměr ve výčetní výšce v cm měřený na základě obvodu; DEF - odhad defoliace v procentech
strom číslo |
druh |
výška 1987 |
výška 1993 |
DBH 1987 |
DBH 1989 |
DBH 1993 |
DBH 1997 |
DEF 1993 |
DEF 1994 |
DEF 1997 |
1 |
Q |
24.5 |
23.0 |
19.7 |
19.9 |
21.0 |
21.3 |
18 |
40 |
80 |
2 |
Q |
22.5 |
23.0 |
20.7 |
21.2 |
22.4 |
22.4 |
12 |
45 |
75 |
3 |
Q |
22.5 |
26.0 |
25.2 |
25.5 |
26.5 |
26.7 |
20 |
55 |
60 |
4 |
Q |
22.0 |
25.0 |
26.1 |
26.8 |
27.5 |
27.4 |
18 |
40 |
65 |
5 |
Q |
21.5 |
27.0 |
28.3 |
29.0 |
31.0 |
32.3 |
18 |
40 |
55 |
6 |
Q |
19.0 |
22.0 |
17.2 |
17.5 |
17.9 |
17.8 |
25 |
40 |
70 |
7 |
Q |
19.0 |
26.0 |
24.5 |
25.5 |
26.7 |
27.1 |
37 |
40 |
65 |
8 |
Q |
16.5 |
[-] |
17.2 |
17.0 |
[-] |
[-] |
[-] |
[-] |
[-] |
9 |
Q |
17.0 |
* |
20.4 |
20.5 |
20.9 |
[-] |
100 |
[-] |
[-] |
10 |
Q |
23.0 |
27.0 |
41.1 |
42.7 |
45.8 |
47.6 |
43 |
55 |
45 |
11 |
Q |
16.0 |
* |
14.6 |
14.6 |
15.1 |
[-] |
80 |
[-] |
[-] |
12 |
Q |
20.0 |
23.0 |
21.7 |
22.5 |
23.4 |
23.9 |
17 |
50 |
60 |
13 |
Q |
19.0 |
25.0 |
22.3 |
22.9 |
23.4 |
23.6 |
20 |
45 |
80 |
14 |
Q |
19.0 |
25.0 |
26.8 |
27.1 |
28.8 |
29.9 |
8 |
35 |
45 |
15 |
Q |
19.5 |
25.0 |
22.3 |
22.6 |
23.5 |
23.9 |
18 |
45 |
50 |
16 |
Q |
21.0 |
26.0 |
30.3 |
31.5 |
33.4 |
34.7 |
33 |
60 |
40 |
17 |
Q |
20.0 |
25.0 |
24.5 |
25.0 |
25.8 |
26.1 |
17 |
45 |
65 |
18 |
Q |
18.5 |
21.5 |
19.4 |
20.1 |
20.5 |
20.1 |
70 |
80 |
100 |
19 |
A |
13.0 |
16.5 |
16.6 |
17.5 |
18.0 |
18.8 |
2 |
5 |
5 |
20 |
A |
13.5 |
16.0 |
15.9 |
17.0 |
17.2 |
18.3 |
2 |
5 |
5 |
21 |
Q |
17.0 |
20.5 |
16.9 |
17.4 |
17.7 |
17.5 |
20 |
70 |
100 |
22 |
A |
16.0 |
17.5 |
20.1 |
21.5 |
22.0 |
22.6 |
0 |
5 |
5 |
23 |
Q |
19.0 |
25.0 |
26.1 |
26.6 |
27.6 |
27.9 |
33 |
35 |
40 |
24 |
Q |
18.0 |
[-] |
17.5 |
17.4 |
[-] |
[-] |
[-] |
[-] |
[-] |
25 |
Q |
22.0 |
26.5 |
33.1 |
34.1 |
35.6 |
36.6 |
25 |
35 |
40 |
26 |
Q |
14.5 |
* |
15.0 |
14.8 |
14.7 |
[-] |
60 |
100 |
[-] |
27 |
Q |
17.5 |
19.0 |
16.9 |
16.2 |
17.4 |
16.9 |
28 |
45 |
55 |
28 |
Q |
17.0 |
* |
15.0 |
15.0 |
15.3 |
15.3 |
30 |
70 |
95 |
29 |
Q |
21.0 |
24.5 |
25.5 |
26.1 |
26.9 |
27.1 |
23 |
30 |
40 |
30 |
Q |
20.5 |
25.5 |
28.0 |
29.3 |
30.4 |
31.0 |
53 |
45 |
65 |
31 |
Q |
18.0 |
22.5 |
18.5 |
18.8 |
19.5 |
19. |
22 |
35 |
30 |
32 |
Q |
19.0 |
25.0 |
20.4 |
21.3 |
22.2 |
22.9 |
25 |
55 |
40 |
33 |
Q |
18.0 |
23.0 |
21.7 |
22.0 |
22.6 |
22.6 |
35 |
55 |
40 |
34 |
Q |
17.0 |
[-] |
18.2 |
17.5 |
[-] |
[-] |
[-] |
[-] |
[-] |
35 |
Q |
19.5 |
25.0 |
23.6 |
24.8 |
26.2 |
27.1 |
25 |
40 |
35 |
36 |
Q |
* |
23.0 |
* |
21.7 |
22.5 |
[-] |
28 |
40 |
[-] |
37 |
Q |
* |
24.5 |
* |
28.2 |
29.5 |
30.2 |
40 |
35 |
35 |
38 |
Q |
* |
* |
* |
19.7 |
19.5 |
19.1 |
100 |
100 |
100 |
39 |
Q |
* |
* |
* |
18.0 |
18.2 |
[-] |
100 |
100 |
[-] |
[-] - strom byl vytěžen po jeho odumření nebo jako velmi silně poškozený
* - parametr nebyl stanoven
průměr |
20.0 |
24.3 |
23.7 |
24.3 |
25.3 |
25.7 |
30 |
49 |
60 |
směr.odchylka |
1.9 |
2.0 |
5.6 |
5.8 |
6.3 |
6.8 |
19 |
15 |
21 |
n |
26 |
26 |
26 |
28 |
28 |
28 |
28 |
28 |
28 |
Poznámka: V úvahu byly brány pouze duby, které nebyly vytěženy do doku 1997.
Back to the main page IDS
© Karel Matějka - IDS (2003)